




《猫猫钓游记》可爱+收集+钓鱼游戏试玩
2026-06-30
2026-07-02 0
DRUGONE
AlphaFold3 能够从序列预测高精度蛋白质结构,但通常会收敛到单一主导构象,即使真实结构本身具有明显异质性,也难以充分呈现构象集合。此外,AlphaFold3 的预测通常不会考虑实验条件,而实验条件可能改变局部序列构象。研究人员提出一种实验引导的 AlphaFold3 方法,使其能够匹配核磁共振、X 射线晶体学和冷冻电镜实验数据,也可以同时整合多种实验模态。该方法还能够纳入直接反映动力学的信息,例如位点分辨的有序参数。结果显示,该方法可以生成紧凑的蛋白质结构构象集合,其集合平均观测量与实验结果一致,相比传统核磁共振解析结构具有更少的距离约束违背,并能在电子密度中发现此前未建模的替代构象。该研究为“实验感知”的预测模型提供了新方向,使模型能够生成与多模态测量结果一致的结构集合,并有望进一步结合能量信息,得到更具热力学基础的构象集合。

蛋白质本质上是动态分子,会根据环境变化和生物学功能需求在多个构象状态之间采样。X 射线晶体学、核磁共振和冷冻电镜等实验技术通常观测到的是大分子结构的集合平均信号,但结构模型往往只报告一个主导构象,从而忽略真实存在的构象异质性。
AlphaFold 系列模型的成功建立在蛋白质空间相邻氨基酸具有共同进化信号这一基础发现之上,并通过大量实验解析结构训练,实现了接近实验精度的结构预测。然而,AlphaFold 的训练目标主要是预测一个“最可能”的结构,这使其输出偏向静态快照,难以显式表达训练数据中蕴含的构象异质性。因此,结构生物学需要新的模型框架,能够生成与实验测量一致的蛋白质结构集合。
研究人员将 AlphaFold3 重新理解为一种“序列条件结构先验”,并把构象集合建模表述为在给定实验测量条件下的后验结构推断问题。与以往将实验约束加入单一结构预测的方法不同,本研究直接修改 AlphaFold3 的生成过程,在采样过程中引入实验似然作为引导项,使模型能够针对任意实验模态生成同时符合序列信息和实验数据的构象集合。
方法
研究人员通过修改 AlphaFold3 反向扩散采样步骤,使其包含由实验似然函数导出的梯度引导项。该项相当于一种数据依赖的“力”,在采样过程中推动结构朝向与实验测量相容的方向移动。一个缩放超参数用于控制实验引导强度,使模型可以在完全依赖 AlphaFold3 先验和强实验约束之间进行调节。该框架可通过定义可微分似然函数支持多种实验数据,包括核磁共振距离约束、有序参数、X 射线电子密度图和冷冻电镜静电势图。采样完成后,研究人员进行力场能量最小化,以修正几何畸变并保证化学合理性;随后通过集合筛选选择能够最好解释实验观测的最小结构子集。
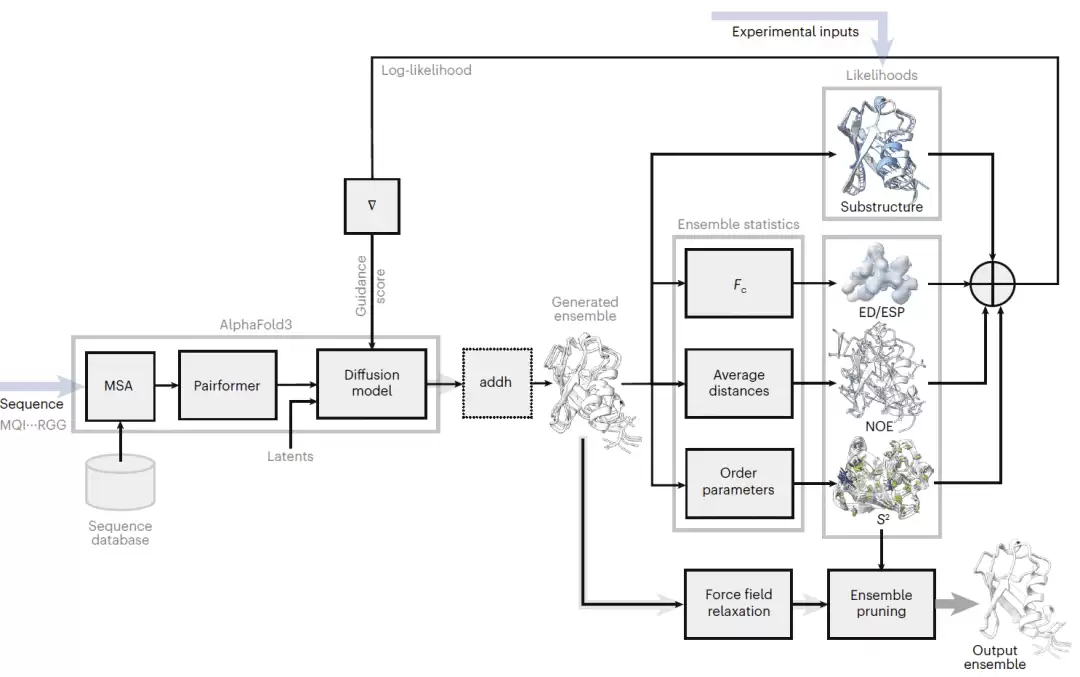
图1|实验引导 AlphaFold3 方法示意图。
结果
实验引导 AlphaFold3 生成蛋白质构象集合
研究人员将 AlphaFold3 扩展为能够生成构象集合的模型。在每一个反向扩散时间步,模型不再只生成单一结构,而是生成一组结构,并根据实验观测计算似然。实验似然既可以作用于单个构象,也可以作用于集合平均量,例如平均原子间距离、电子密度、有序参数等。随后,实验似然的梯度作为引导项反馈到扩散采样过程中。最终得到的构象集合经过力场松弛和集合剪枝,保留最能解释实验数据的紧凑结构集合。
核磁共振距离约束和有序参数引导 AlphaFold3
研究人员首先以泛素作为核磁共振结构解析基准。传统核磁共振结构解析主要依赖原子间距离约束,但逐个模拟构象容易产生模式坍缩,得到过于刚性的集合,无法充分反映真实动力学。相比之下,NOE 距离约束引导的 AlphaFold3 能够生成更符合实验距离约束的构象集合,并显著减少距离约束违背。与传统 PDB 核磁共振集合和未引导的 AlphaFold3 相比,NOE 引导模型产生的集合更加异质,计算时间也仅需数分钟 GPU 时间,从而避免了传统集合分子动力学方法的高计算成本。
研究人员进一步使用主链酰胺键有序参数进行交叉验证。未引导 AlphaFold3 和传统核磁共振集合均偏向刚性构象,与实验动力学信号的一致性有限。NOE 引导 AlphaFold3 能够更好重现实验观测到的柔性区域和结构区域动力学。当进一步加入有序参数引导项后,模型与实验有序参数的匹配程度显著提高,并接近计算成本更高的核磁共振引导集合分子动力学结果。
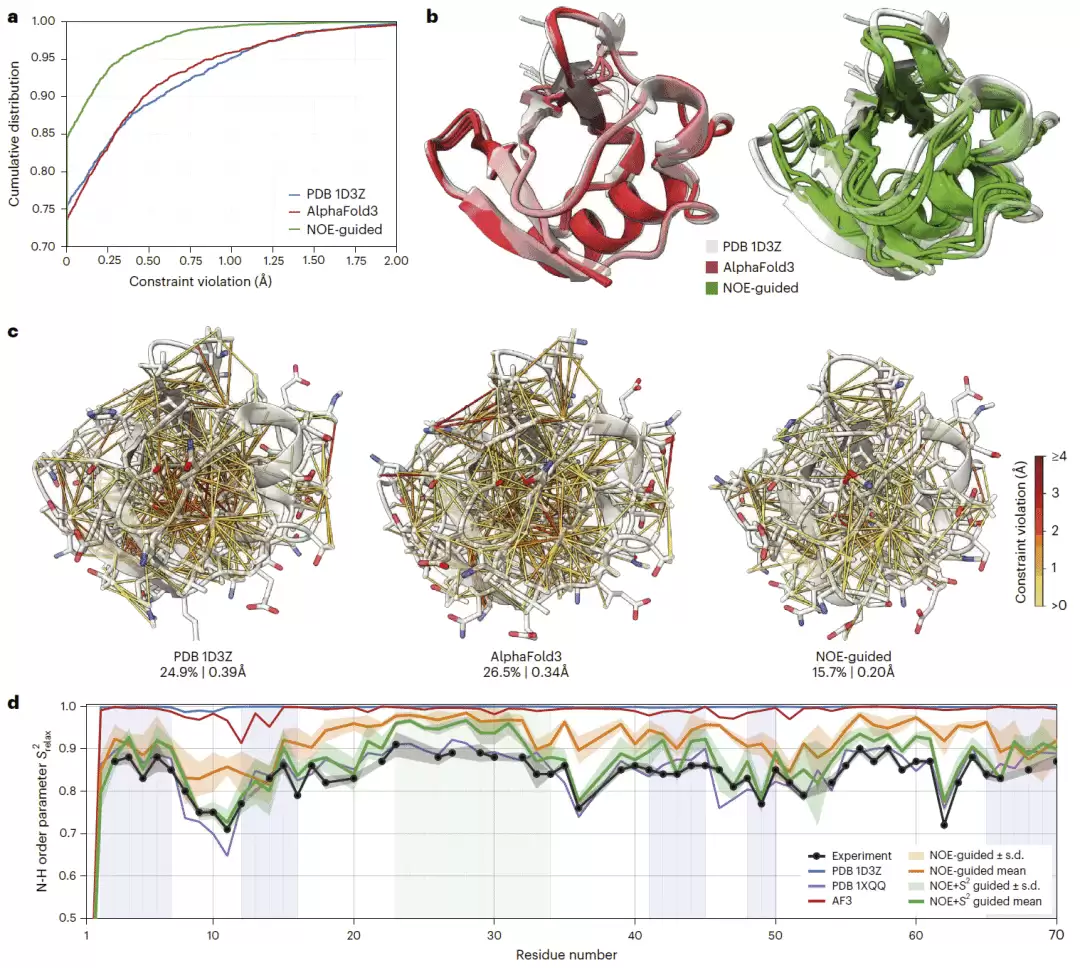
图2|核磁共振引导的 AlphaFold3。
核磁共振基准测试
研究人员进一步在两个基准上评估该方法,包括此前已知 AlphaFold3 容易预测错误的8个肽结构,以及一个包含83个蛋白质的核磁共振谱数据库子集。总体结果显示,与 PDB 中沉积的构象集合相比,NOE 引导 AlphaFold3 在91个案例中的70个案例改善了距离约束满足程度,比例约为77%;而未引导 AlphaFold3 仅在15个案例中优于 PDB 集合。NOE 引导 AlphaFold3 在所有案例中均优于未引导 AlphaFold3。
研究人员还测试了将力场预测能量用于构象权重调整,或将力场直接整合进引导项。后一种方式进一步提高了距离约束满足度,在约87%的案例中取得改善,中位改善幅度约为20%。这说明实验约束和能量信息可以互补,有助于将采样分布推向更具热力学意义的构象集合。
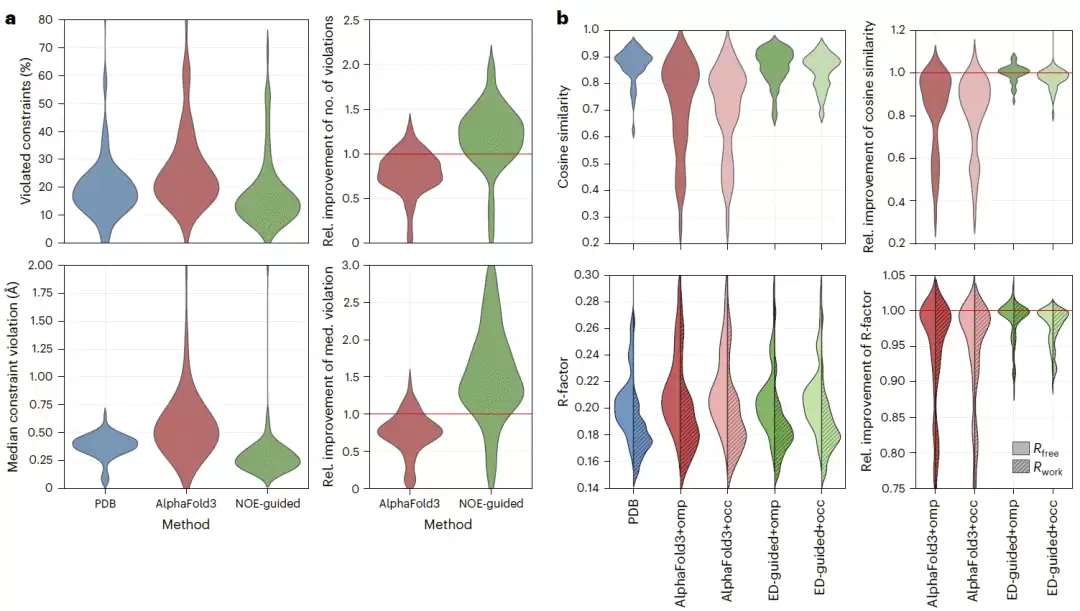
图3|核磁共振和 X 射线晶体学基准测试的总体统计。
六个蛋白质构象集合案例
在具体案例中,未引导 AlphaFold3 往往过度预测有序结构,例如生成过长的螺旋片段。NOE 引导能够增加集合异质性,改善距离约束满足,并在多个案例中优于相应 PDB 核磁共振集合。有些蛋白质中,AlphaFold3 对局部结构完全预测错误,导致大量距离约束违背,而 NOE 引导可明显减少这些违背。对于某些结构,没有任何单一构象能完全满足实验距离约束,但集合平均引导能够更好解释实验数据,这凸显了“构象集合”而非“单一结构”在解释实验测量中的必要性。
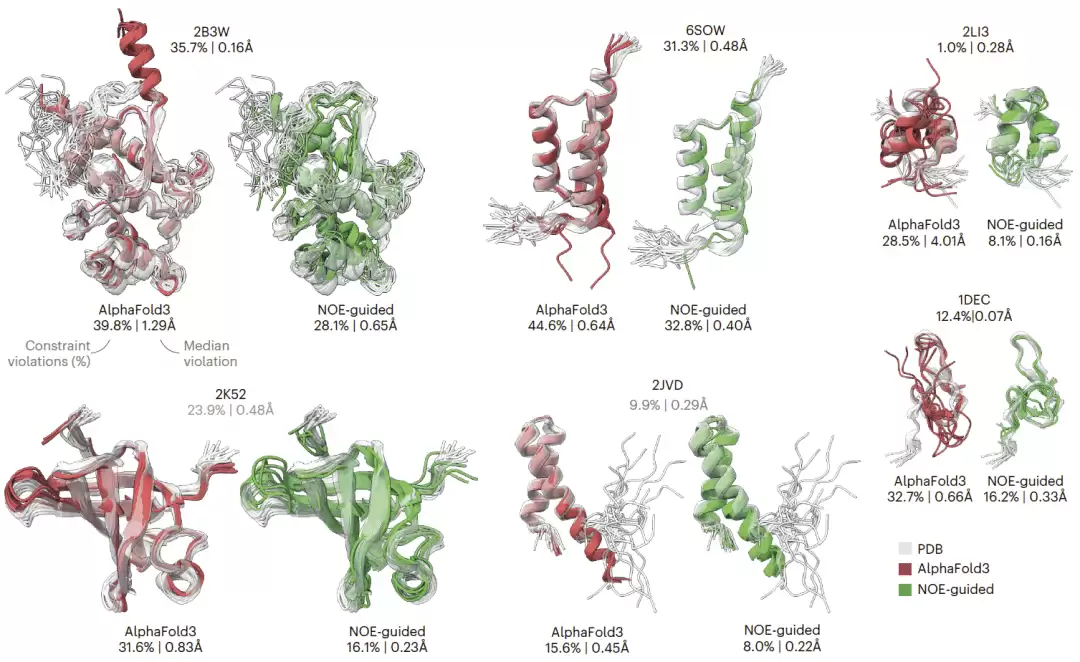
图4|六个蛋白质的构象集合生成示例。
电子密度引导 AlphaFold3 捕获 X 射线晶体学中的未建模构象
AlphaFold3 通常不会感知晶体环境中的离子、配体、其他大分子伙伴或晶体接触等因素,而这些因素可能导致局部构象变化。研究人员显示,电子密度引导可以将 AlphaFold3 预测修正到与晶体结构实验相符的状态。例如,在 HSP90α 中,配体结合状态影响邻近环区构象,未引导 AlphaFold3 无法准确预测,而电子密度引导能够恢复与实验一致的局部结构。
该方法还可处理分辨率不同的电子密度图,并能在不同蛋白、不同长度序列和不同晶体环境中重建实验观测到的替代构象。对于晶体结构中因局部柔性而缺失的区域,电子密度引导 AlphaFold3 能够根据稀疏密度填补缺失片段,并比普通 AlphaFold3 或常用修补工具更好解释局部电子密度。对于短肽结构,AlphaFold3 往往预测较差,因为短肽高度依赖结合伙伴环境;电子密度引导可以显著恢复肽段与实验密度的一致性。
研究人员还展示了该方法捕获晶体结构中替代位置的能力。即使对于双峰或多峰电子密度,电子密度引导 AlphaFold3 也能生成能够解释多模态密度的构象集合,并有潜力发现此前未建模的替代构象。
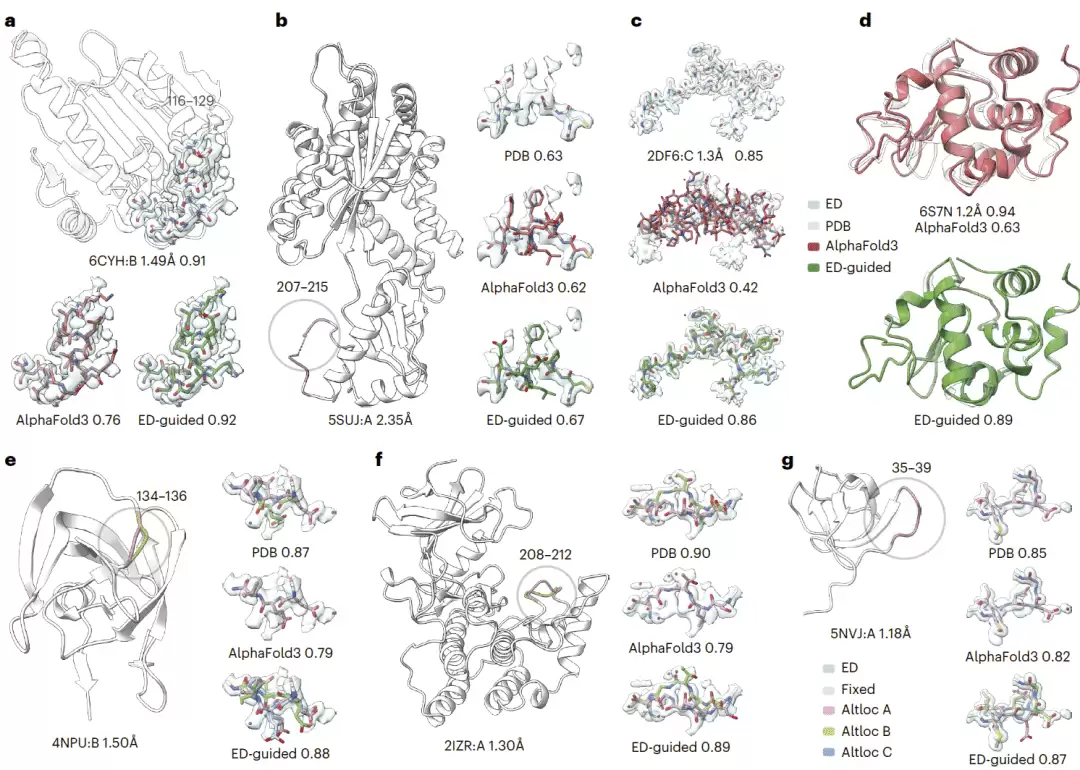
图5|X 射线电子密度引导 AlphaFold3 重建结构和构象集合。
冷冻电镜图引导 AlphaFold3
冷冻电镜可以通过静电势图捕获复杂蛋白复合物的构象,但标准重构流程往往依赖刚体共识模型,单个静电势图难以完整呈现粒子的真实热力学异质性。同时,柔性区域的局部分辨率常不足以精确放置每个原子。研究人员在多个案例中显示,使用冷冻电镜静电势图引导 AlphaFold3 可以提高从头建模精度。
在胰岛素受体 IR-B 的对称 apo 状态和结合 IGF2 的非对称构象中,AlphaFold3 倾向于过度对称化同源二聚体,尤其会错误预测非对称构象的大部分区域。静电势图引导后,模型生成的构象更符合观测密度。在淀粉样 β 纤维案例中,AlphaFold3 完全误判二聚化界面和二聚单元结构,而静电势图引导能够使结构与密度图更好一致。
联合冷冻电镜和核磁共振数据引导 AlphaFold3
结构生物学研究常常同时使用多种实验技术。该框架可以整合冷冻电镜、核磁共振等多源数据。研究人员以 RIPK3 人源淀粉样纤维为例,联合使用冷冻电镜静电势图和固态核磁共振化学位移推断的二面角信息。AlphaFold3 本身可以在冷冻电镜分辨率范围内较好预测整体多聚体结构,单独使用静电势图引导可提高密度拟合质量,但对局部主链二面角和 NOE 约束拟合较差。联合使用静电势图、NOE 和二面角引导后,模型在保持合理密度拟合的同时,提高了局部结构准确性。
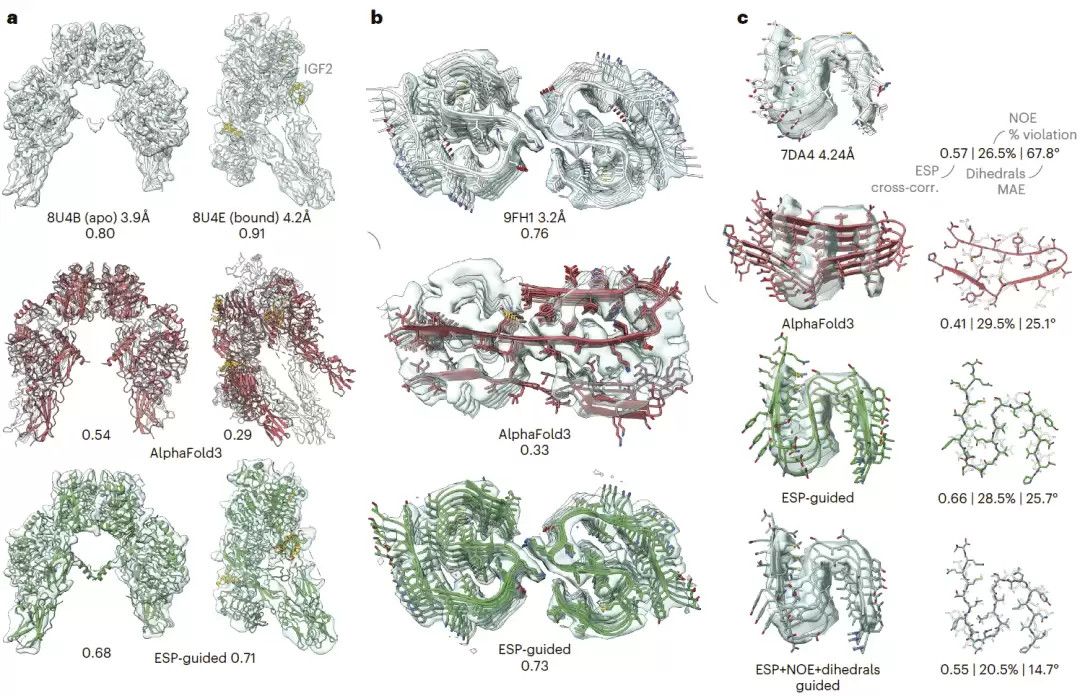
图6|冷冻电镜静电势图引导的多聚体结构重建。
讨论
本研究将 AlphaFold3 重新定义为一种强大的序列条件先验模型,它可以被实验数据引导,从而生成小型、可检验的蛋白质构象集合,而不是单一共识结构。这些集合应被理解为受实验测量和模型先验共同约束的后验结构假设,而不是已经完全校准的玻尔兹曼平衡集合。换言之,该方法解决的是结构生物学中一个非常实际的问题:生成一组紧凑构象,使其集合平均观测量能够解释异质实验数据。
该框架的核心优势在于,不同实验模态的似然函数可以组合,包括核磁共振原子间距离、二面角、有序参数,X 射线晶体学电子密度,以及冷冻电镜静电势图。这些实验信息通过梯度注入 AlphaFold3 的反向扩散轨迹,再经过结构松弛和稀疏集合选择,最终得到可解释实验数据的构象集合。与传统 PDB 表示方式相比,这种方法能够更充分呈现蛋白质构象景观中被单一结构或简化集合所掩盖的信息。
方法学上,本研究有三个关键特点。第一,构象集合在扩散过程中被联合引导,因此在实验数据允许多种解释时可以保留多样性,同时避免简单复制同一构象。第二,该框架支持从局部到整体的策略,可以先针对环区等问题片段进行修正,再扩展到整条蛋白链,这与晶体学和核磁共振研究人员的实际工作方式相似。第三,计算成本较低,典型案例仅需数分钟,因此更适合作为现有结构解析流程中的交互式助手,而不是完全替代精修或分子模拟。
不过,研究人员也强调了若干限制。该方法生成的构象集合主要是对给定实验数据和 AlphaFold3 先验的解释性集合,并不等同于真实热力学平衡群体。有限实验观测不能唯一确定底层构象分布,不同构象集合可能同样解释数据。因此,集合中不同构象的权重不应被过度解读为平衡态群体比例。加入力场能量后结果进一步改善,说明能量信息提供了与实验约束互补的限制,未来需要更紧密整合能量先验和热力学观测。
此外,引导强度和梯度处理也非常重要,尤其是在低分辨率冷冻电镜图或相位较差的晶体学电子密度图中,过强引导可能把模型拉向噪声。虽然集合剪枝和交叉验证可以缓解过拟合,但未来仍需要更严格的统计程序来区分真实热力学熵和模型不确定性。当前实现主要聚焦蛋白质,对配体、金属、翻译后修饰、结构水和复杂对称性的处理仍有限。更准确地建模配体也将有助于区分天然热力学构象异质性和组成异质性。
总体来看,实验引导 AlphaFold3 为主流结构生物学流程提供了新的改进路径。在晶体学中,它可以帮助提出替代构象和占有率,修复弱电子密度区域中的环区,并通过提供少量已符合密度图的候选构象加速模型构建。在核磁共振中,它可以快速解释 NOESY 谱和弛豫数据,而不需要耗时数天的分子动力学模拟。在冷冻电镜中,它可以将原子级细节注入柔性区域,并在局部分辨率有限时结合二面角或二级结构信息。研究人员认为,实验引导的生成模型有望让以构象集合为中心的结构建模成为晶体学、核磁共振和冷冻电镜中的常规工具,并在配体发现、变异解释以及稳定或揭示隐蔽构象状态的实验设计中产生近期影响。
整理 | DrugOne团队
参考资料
Maddipatla, A., Sellam Bojan, N., Bojan, M. et al. Experiment-guided AlphaFold3 resolves measurement-consistent protein ensembles. Nat Biotechnol (2026).
https://doi.org/10.1038/s41587-026-03166-5
内容为【DrugOne】公众号原创|转载请注明来源
本文参与腾讯云自媒体同步曝光计划,分享自微信公众号。原始发表:2026-06-30,如有侵权请联系[email protected] 删除Copyright(C) 2020-2026 jiyx.com All Rights Reserved 联系方式:[email protected]